脂肪酸 是一元羧酸的组别名称,即由一个组成的化合物 羧基 (-COOH),由不同长度但几乎完全是无支链的烃链组成。命名脂肪酸一方面是由于这组物质的化学性质,由于它们的羧基而具有酸性反应。另一方面,这个名字可以追溯到发现这些烷烃羧酸作为天然脂肪成分的历史。从这个角度来看,脂肪酸被计入脂质中。
一般
脂肪酸的碳原子数(链长)和双键的可能存在、数量和位置不同。脂肪酸可根据其链长分为低级 (最多七个碳原子),中等 (八到十二个碳原子)和更高的 (超过十二个碳原子)脂肪酸。作为脂肪酸的历史命名表明,为了成为脂肪酸,单个化合物必须是脂肪的组成部分。但情况不一定如此。化学相似的化合物现归于该术语。
天然脂肪酸通常由偶数个碳原子组成并且是无支链的。碳链必须至少有四个碳原子长,使丁酸成为最简单的天然脂肪酸。带有双键的脂肪酸称为不饱和脂肪酸。双键通常具有顺式构型。如果有多个双键,它们通常用 CH2 分隔 -分组分开。
在植物界的种子油中发现了各种各样的脂肪酸(超过 400 种不同的结构,但只有大约 10-12 种是常见的)。在某些植物科的种子中大量存在的稀有脂肪酸可以说明发育关系(关系、化学分类学、进化;参见例如千岁)。
必需脂肪酸是人体无法从其他营养素合成的脂肪酸。所有那些在第九个碳原子的末端(远离羧基)至少有一个双键的脂肪酸都是人体必需的。
高级脂肪酸的钠盐或钾盐称为皂类,用作表面活性剂。
它们主要在食品工业中用作各种乳化剂的原料,也用作载体、脱模剂(例如口香糖)或涂层(例如水果)。在欧盟,它们被归类为食品添加剂统称 E 570 没有最大限制 (quantum satis) 一般批准用于食品。
饱和和不饱和脂肪酸
A 饱和脂肪酸 是一种在碳原子之间没有双键的脂肪酸。饱和脂肪酸形成分子式Cn的所谓同源序列 H2n+1 二氧化碳。 不饱和脂肪酸 有一个双键和多不饱和脂肪酸 链的碳原子之间有两个或多个双键。由于天然脂肪酸中的双键多为顺式构型,因此烃链中存在约30°的扭结。这削弱了与其他分子的范德华相互作用并降低了熔点。一些不饱和脂肪酸对人类来说是必需的(生命必需的),因为它们不能被人体合成。这些包括在某些位置带有双键的脂肪酸,即所谓的 omega-n 脂肪酸 .这里 n 代表一个数字并描述了其中一个双键的位置。使用 omega 计数 (ω-),从 结束 从碳链开始计算,羧基形成分子的开头。如果有多个双键,则只有第一个双键的位置很重要。在亚麻酸的插图中,ω 计数以红色显示。 (另请参阅:Omega-3、Omega-6 和 Omega-9 脂肪酸)。除了顺式构型的不饱和脂肪酸外,自然界中也存在反式构型的脂肪酸,即所谓的反式脂肪酸 .反式脂肪酸也是人造黄油生产的副产品,被怀疑具有损害健康的特性。如果脂肪酸中有多个双键,这些双键通常 - 类似于右上方所示的亚麻酸 - 通过 CH2 -组彼此分开。然而,也有所谓的共轭脂肪酸 ,其中双键更靠近,即共轭。在 octadeca-9c,11t-二烯酸(右)的图片中,双键是共轭的。由于其中一个双键是反式双键,因此该化合物也是反式脂肪酸。反刍动物消化道中的细菌通常是这些脂肪酸形成的原因。因此,共轭脂肪酸存在于所有乳制品中。
支链一元羧酸
支链羧酸通常不计入脂肪酸,但存在于一些精油中。缬草提取物含有异戊酸酯。
植烷酸(3,7,11,15-四甲基十六烷酸)是一种支链羧酸,作为叶绿素的分解产物出现。这种化合物的痕迹可以在许多食物(例如牛奶)中找到。病理性不能分解这种羧酸导致Refsum综合征。
脂肪酸和较短的一元羧酸列表
| 饱和脂肪酸和一元羧酸 | ||||||
|---|---|---|---|---|---|---|
| 碳原子数:双 键 | 平凡的名字 | 总公式 | 出现次数 | 熔点 点 | 化学名称 | |
| 1:0 | 甲酸 | HCOOH | 广泛存在于几乎所有生物体中 | 8°C | 甲酸 | |
| 2:0 | 醋酸 | CH3 二氧化碳 | 醋(通过乙醇氧化) | 16.2°C | 乙酸 | |
| 3:0 | 丙酸 | C2 H5 二氧化碳 | 甲烷发酵中间产物 | -24 °C | 丙酸 | |
| 4:0 | 丁酸 | C3 H7 二氧化碳 | 乳脂,汗水 | -8 °C | 丁酸 | |
| 5:0 | 戊酸 | C4 H9 二氧化碳 | 缬草、木醋液 | -35 °C | 戊酸 | |
| 6:0 | 己酸 | C5 H11 二氧化碳 | 丁酸发酵过程中产生的乳脂 | -4 °C | 己酸 | |
| 7:0 | 庚酸 | C6 H13 二氧化碳 | 作为菖蒲油中的酯 | -7.5°C | 庚酸 | |
| 8:0 | 辛酸 | C7 H15 二氧化碳 | 乳脂、椰子油 | 17°C | 辛酸 | |
| 9:0 | 壬酸 | C8 H17 二氧化碳 | 玫瑰天竺葵精油、奶酪、杂醇油、葡萄酒 | 12.5°C | 壬酸 | |
| 10:0 | 癸酸 | C9 H19 二氧化碳 | 动植物脂肪 | 31°C | 癸酸 | |
| 12:0 | 月桂酸 | C11 H23 二氧化碳 | 乳脂、植物脂肪 | 43.2°C | 十二烷酸 | |
| 14:0 | 肉豆蔻酸 | C13 H27 二氧化碳 | 乳脂、鱼油、动植物脂肪 | 53.9°C | 十四烷酸 | |
| 16:0 | 棕榈酸 | C15 H31 二氧化碳 | 动植物脂肪 | 62.8°C | 十六烷酸 | |
| 17:0 | 柠檬酸 | C16 H33 二氧化碳 | 动植物脂肪 | 61.3°C | 十七烷酸 | |
| 18:0 | 硬脂酸 | C17 H35 二氧化碳 | 动植物脂肪 | 69.6°C | 十八烷酸 | |
| 20:0 | 花生酸 | C19 H39 二氧化碳 | 少量存在于植物种子和动物脂肪中 | 75.4°C | 二十烷酸/二十烷酸 | |
| 22:0 | 山嵛酸 | C21 H43 醇 | 少量存在于植物种子和动物脂肪中,导致戈谢病 | 二十二烷酸 | ||
| 24:0 | 木蜡酸 | C23 H47 醇 | 一些植物脂肪,部分鞘磷脂 | 十四烷酸 | ||
| 26:0 | 蜡酸 | C25 H51 醇 | 蜂蜡、巴西棕榈蜡、褐煤蜡、羊毛汗水 | 十六烷酸 | ||
| 单不饱和脂肪酸 | ||||||
|---|---|---|---|---|---|---|
| 碳原子数:双 键 | 平凡的名字 | 总公式 | 位置 | 出现次数 | 熔点 点 | 化学名称 |
| 11:1 | 十一碳烯酸 | C10 H19 二氧化碳 | 10 | 24.5°C | (10Z )-undeca-10-enoic acid | |
| 16:1 | 棕榈油酸 | C15 H29 二氧化碳 | 9 | 乳脂、动物储存脂肪、鱼油、植物脂肪 | (9Z )- 十六- 9-烯酸 | |
| 18:1 | 油酸 | C17 H33 二氧化碳 | 9 | 在所有天然脂肪中 | 16℃ | (9Z )- octadeca- 9-enoic acid |
| 18:1 | 反油酸 | C17 H33 二氧化碳 | 9 | 在反刍动物脂肪中 | 44-51°C | (9E )- octadeca- 9-enoic acid |
| 18:1 | 异油酸 | C17 H33 二氧化碳 | 11 | (11E )- octadeca- 11-enoic acid | ||
| 20:1 | 二十碳烯酸 | C19 H37 二氧化碳 | 11 | 在菜籽油中 | 16℃ | (11Z )- 二十碳- 11-烯酸 |
| 22:1 | 鲸蜡酸 | C21 H41 二氧化碳 | 11 | 植物油 | (11Z )- 二十二烷- 11-烯酸 | |
| 22:1 | 芥酸 | C21 H41 二氧化碳 | 13 | 菜籽油 | (13Z )- 二十二烷- 13-烯酸 | |
| 24:1 | 神经酸 | C23 H47 二氧化碳 | 15 | (15Z )- 二十碳糖- 15-烯酸 | ||
| 多不饱和脂肪酸 | ||||||
|---|---|---|---|---|---|---|
| 碳原子数:双 键 | 平凡的名字 | 总公式 | 位置 | 出现次数 | 熔点 点 | 化学名称 |
| 18:2 | 亚油酸 | C17 H31 二氧化碳 | 9, 12 | 植物油,尤其是红花油 | -5 °C | (9Z ,12Z )- octadeca-9,12-二烯酸 |
| 18:3 | 亚麻酸 | C17 H29 二氧化碳 | 9、12、15 | 一些植物油,尤其是亚麻籽油 | -11 °C | (9Z ,12Z ,15Z )- octadeca-9,12,15-三烯酸 |
| 20:4 | 花生四烯酸 | C19 H31 二氧化碳 | 5、8、11、14 | 动物脂肪、鱼油 | -49.5 °C | (5Z ,8Z ,11Z ,14Z )- Eicosa- 5,8,11,14- 四烯酸 |
| 20:5 | 替莫诺宁酸 | C19 H29 二氧化碳 | 5、8、11、14、17 | 鱼油 | -54 °C | (5Z ,8Z ,11Z ,14Z ,17Z )- Eicosa- 5,8,11,14,17-戊烯酸 |
| 22:5 | 鲶鱼酸 | C21 H33 二氧化碳 | 7、10、13、16、19 | 鱼油 | (7Z ,10Z ,13Z ,16Z ,19Z )- Docos- 7,10,13,16,19-戊烯酸 | |
| 22:6 | 鹿茸酸 | C21 H31 二氧化碳 | 4、7、10、13、16、19 | 鱼油 | (4Z ,7Z ,10Z ,13Z ,16Z ,19Z )- Docosa- 4,7,10,13,16,19-六烯酸 | |
注意事项:
- 反油酸,油酸的反式异构体,是在人造黄油生产过程中通过异构化过程中多不饱和脂肪酸的部分氢化而在脂肪硬化过程中形成的。它天然存在于反刍动物(牛奶、黄油、牛油)的脂肪中,因为它们的瘤胃生物也含有氢化酶。
- Vaccenic 酸是一种反式脂肪酸,存在于反刍动物乳制品中。
- 在 9、12 和 15 位具有双键的亚油酸异构体(均为顺式构型)是 α-亚麻酸,即在 6、9 和 12 位具有双键的异构体(均为顺式构型)被称为 γ-亚麻酸。
新陈代谢
脂肪酸分解
脂肪酸以甘油三酯的形式储存在脂肪组织中。如果需要,由信使物质肾上腺素、去甲肾上腺素、胰高血糖素或促肾上腺皮质激素指示,在那里发生脂肪分解。
然后游离脂肪酸在血流中被运输到需要能量的细胞,在那里它们首先与辅酶 A (CoA) 结合(活化),同时消耗 ATP。该反应通过生成的焦磷酸盐水解成两种磷酸盐(Pi )。
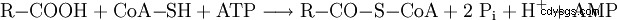
然后它们通过肉碱酰基转移酶 I 与肉碱结合并转运到线粒体基质,在此它们通过肉碱酰基转移酶 II 与 CoA 结合。
在线粒体基质中,脂肪酸发生β-氧化为乙酰辅酶A,可进一步用于柠檬酸循环以获得ATP。在长期饥饿或碳水化合物极少的不均衡饮食期间,例如B.阿特金斯饮食法,脂肪被代谢成酮体。
除了线粒体脂肪酸氧化外,脂肪酸利用也发生在过氧化物酶体中。特别是非常长的链脂肪酸通常首先在那里被缩短,然后才能在线粒体中进一步加工。这种过氧化物酶体功能很重要。失败会导致肾上腺脑白质营养不良。
脂肪酸合成
与降解相反,脂肪酸合成发生在胞质溶胶中。在高等生物中,所有必需的酶都结合在一个酶复合物中,即脂肪酸合酶。然而,在绿色植物中,C18脂肪酸的积累主要发生在质体中,然后才被转运到细胞质中。
为此目的,丙二酰辅酶A首先由乙酰辅酶A通过羧化并消耗ATP形成。然后将其转化为丙二酰-ACP,因为与降解相比,酰基载体蛋白 (ACP) 在合成过程中充当载体分子而不是 CoA。随后的缩合反应大致是脂肪酸氧化(β-氧化)的逆转。但是,在细节上存在一些显着差异,允许对两个过程进行独立、有针对性的控制。
健康重要性
饱和脂肪提供大量能量,但不是必需的。中链脂肪酸含量高的脂肪比长链脂肪酸含量高的脂肪更容易消化。反式脂肪酸对胆固醇水平和心脏病发作的风险有不利影响。 Omega-6 脂肪酸被代谢为促进炎症的前列腺素,而 omega-3 脂肪酸则为抗炎脂肪酸。
DGE 建议用脂肪覆盖大约 30% 的能量需求。 10% 应覆盖饱和脂肪,10 至 13% 覆盖单不饱和脂肪,其余为多不饱和脂肪。为了降低心血管风险,omega-6与omega-3脂肪酸的比例不应超过5:1。
- 脂质
- 脂肪酸合成
- Omega-3 脂肪酸
- Omega-6 脂肪酸
- Omega-9 脂肪酸
来源
- ↑ 参见种子脂肪酸数据库,https://sofa.bfel.de/
- ↑ 联邦营养与食品研究中心
参考文献
- W. H. Kunau:不饱和脂肪酸的化学和生物化学 . Angewandte Chemie 88, 97 (1976)
- J. Ernst, W.S.谢尔德里克和 J.-H。 Fuhrhop:必需不饱和脂肪酸的结构。亚油酸的晶体结构及亚麻酸和花生四烯酸晶体结构的证明 . Z.自然。 1979, 34b, 706-711
- P. Nuhn、M. Gutheil、B. Dobner:支链脂肪酸的发生、生物合成和重要性。 Fette-Seifen-Anstrichmittel 87, 135 (1985)